Dans le prolongement de l’édito consacré à un photogramme de L’Hippocampe ou « cheval marin » (1934) de Jean Painlevé, scientifique proche des surréalistes, j’ai proposé à deux artistes-chercheurs, Jeremie Brugidou et Lia Giraud, dont les travaux créent des images inédites avec des organismes sous-marins, d’explorer les nouvelles visions du vivant. Dans une dialectique arts-sciences, l’un et l’autre bénéficient d’une carte blanche associant théorie et pratique qui aboutira à la réalisation commune pour PALM d’un film prospectif. Jeremie Brugidou décline ses recherches dans de multiples formats. Né en 1988, ancien élève de l’École normale supérieure, il est l’auteur d’articles, de films, du roman Ici, la Béringie (Éditions de l’ogre, 2021) et de la thèse Vers une écologie de l’apparition à paraître cette année aux éditions Mimésis. Actuellement en résidence à l’IMéRA (Institut Méditerranéen de Recherches Avancées / Aix Marseille Université), il prépare pour le 8 juin prochain, Journée mondiale des océans, une installation sous-marine fondée sur la bioluminescence d’une bactérie. Il s’en explique dans « Photogénies transpèces », sa première contribution à PALM qui contextualise son projet. Étienne Hatt
Un œil au fond de la Méditerranée à 2200 mètres. L’obscurité est épaisse, ponctuée de flashs bleus épars et intermittents. À la fin du 19e siècle, ce milieu de vie au volume colossal (plus de 95% de la zone habitable sur Terre) n’existe pas : il n’a pas été mesuré, il n’a pas été sondé ; pour la pensée occidentale cette zone est sans fond et sans vie. À partir du 20e siècle ce volume existe et l’humain·e y est présent·e, par prolongements techniques et médiations épistémologiques : sondes, sonars, submersibles, robots téléguidés, prélèvements de sédiments et de micro-organismes… Ici, en Méditerranée à 2200 mètres de fond, il s’agit d’un observatoire de grande profondeur, initialement dénommé ANTARES, constitué d’1 km3 de globes ultra-sensibles braqués vers le fond de la mer. Cette profondeur extrême existe maintenant en tant que biotope, milieu de vie, mais aussi en tant que poste d’observation, de prélèvement et potentiellement d’exploitation de ressource. Cette zone est nommée le benthos. Elle reste malgré tout faiblement peuplée, c’est la théorie, c’est-à-dire qu’il devrait y faire radicalement noir. Cette obscurité absolue est importante : elle a été choisie pour accueillir des yeux d’un nouveau type, des yeux capables de voir l’invisible. Des globes optiques hypersensibles sont installés au fond de la mer pour former un gigantesque télescope d’1 km3. C’est un invisible très particulier qui est visé ici, un invisible lui aussi absolu, un invisible qu’aucune forme vivante ne peut percevoir, qu’aucun objet technique ne peut saisir directement, un invisible intangible qui traverse la matière en lignes droites, parfaitement mathématiques mais indifférentes au sensible. Je parle d’absolu pour désigner une chose conceptualisée en dehors de tout contexte et qui est donc posée comme immuable, infinie, indifférente.
Au fond de la Méditerranée à 2200 mètres, un œil est posé pour percevoir des particules absolues dans un noir absolu. En effet, selon la théorie, ce noir absolu peut néanmoins à certaines conditions très spéciales être déchiré par la particule absolue. Cette dernière, habituellement indifférente à la matière, peut dans certaines circonstances exceptionnelles s’abaisser à l’interaction mondaine en émettant une autre particule, appelée muon, beaucoup moins indifférente au sensible puisqu’elle peut jouer avec une de ses limites fondamentales : la vitesse de la lumière, qui est alors ralentie par le volume d’eau à forte pression. Un flash bleu est émis, que seul l’œil ultra-sensible du télescope sous-marin peut percevoir : ce flash bleu est produit par un muon lorsqu’il dépasse la vitesse de la lumière dans l’eau (effet Tcherenkov) trahissant alors la présence d’un neutrino à forte énergie, cette fameuse particule absolue que traquent les astronomes depuis le fond de la Méditerranée pour étudier les trous noirs et les formations de galaxies.
Rapidement pourtant ces yeux à neutrinos sont mis hors service. Dans ce volume choisi pour son obscurité absolue, la matière joue avec la matière et le vivant a d’autres projets que ceux des humain·es. Les yeux ultra-sensibles sont rapidement aveuglés, éblouis par des niveaux d’intensité lumineuse inimaginable à de telles profondeurs. Des flashs de lumière surgissent à des niveaux beaucoup plus intenses que ceux produits par les neutrinos et certains yeux se recouvrent d’un voile de lumière persistant. Des globes sont remontés et les biologistes y découvrent une fine pellicule de matière organique déposée sur les lentilles : une colonie bactérienne qui émet de la lumière. Le fond de l’eau, même au plus profond, est parsemé de lumières. Le vivant y passe en nombre, notamment par l’intermédiaire des courants de convection d’eau profonde formés par le refroidissement saisonnier des eaux de surface créant ainsi des circuits de renouvellement des fonds par les surfaces riches en matière organique et des surfaces par les fonds riches en minéraux. Des télescopes, il a fallu passer aux microscopes, de l’infiniment lointain à l’infiniment proche, pour découvrir une nouvelle espèce bactérienne, nommée par les biologistes, Photobacterium phosphoreum ANT-2200.
La vision « absolue » caractérise une perspective autocentrée, ici humaine, vouée tôt ou tard à se heurter aux autres visions émanant du vivant. Une vision détermine donc un champ d’apparition, un domaine circonscrit de possibilité du visible, liée aux capacités motrices, sensorielles et affectives des corps dont elle émane. Une vision n’est cependant pas limitée par la peau, elle peut s’étendre en dehors du corps par prolongements techniques et milieux associés, mais elle mute par transductions et passages de milieu et se différencie au moment où se produit un effet d’échelle suffisamment puissant pour rompre le partage établi entre l’absolu et le relatif.
Je travaille en association avec Photobacterium. Je la cultive et l’utilise, je crée les conditions de possibilités pour qu’une colonie se développe au point de devenir suffisamment lumineuse pour être perceptible par l’œil humain. Bien sûr dans ce processus, une fois le milieu nutritif entièrement consommé et en l’absence d’échappatoire, la colonie dépérit, s’éteint, et meurt. Ensuite le tout est mis à l’autoclave, c’est-à-dire chauffé à 120 degrés, éliminant ainsi toute forme de vie biologique, puis incinéré. Ces bactéries, je les utilise, c’est indéniable. Je tente néanmoins de penser, voire de ménager, la possibilité, si ce n’est d’une réciprocité, au moins d’une correspondance, d’une convergence, même partielle, d’intérêt. Peut-être n’est-il pas possible de penser la moindre forme de réciprocité dans ce processus où j’ai l’impression d’avoir droit de vie et de mort sur ces organismes, aussi unicellulaires soient-ils ; mais si l’on se réfère à des modèles comme la domestication d’un côté ou la physique des forces, par exemple gravitationnelles, d’un autre, on peut postuler qu’il existe toujours un circuit d’influences réciproques, même intangible. On pourrait postuler qu’il existe entre nous l’émission de particules indétectables qui pourtant agissent sur le réel, comme le neutrino, et on pourrait les nommer, à l’inverse du neutrino, le participio. Ainsi, sans jamais oublier le rapport de force qui est en jeu, on peut néanmoins partir d’un principe d’ouverture épistémologique, ou d’empirisme radical, consistant à dire que les bactéries, d’une certaine manière, me cultivent en même temps que je les cultive. La nature de cette manière est un de mes objets d’attention. Nonobstant le déséquilibre flagrant des enjeux entre elles et moi, nous participons ensemble à quelque chose, puisque j’en viens à créer des pouponnières à bactérie. À quel point est-ce que les bactéries me domestiquent à leurs usages et m’utilisent dans tout ce processus reste toutefois à déterminer.
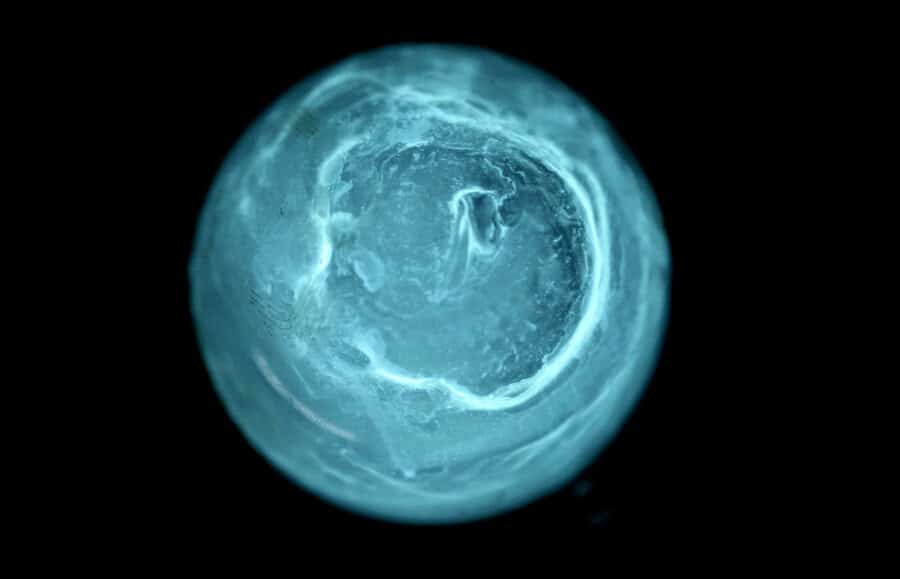
La bioluminescence produite dans les photophores n’est que rarement produite par des bactéries. La symbiose avec bactéries est avérée dans le cas de certaines sépioles, du type Euprymna et Heteroteuthis. La bioluminescence est le plus souvent issue d’une production endogène grâce à une réaction chimique commune à l’ensemble des espèces produisant de la bioluminescence. Néanmoins, cette réaction chimique de base (luciférase+luciférine+oxygène → photon) est peut-être un très ancien héritage bactérien. Visuel © Jeremie Brugidou
On peut tracer quelques grandes lignes. Ces bactéries sont présentes un peu partout dans l’eau de mer, à des concentrations très faibles, et de manière très inégale, par patchs. Elles se développent surtout en grande profondeur, dans des conditions d’obscurité, de basse température et de forte pression sur de la matière organique en suspension charriée par les courants marins. Elles émettent de la lumière pendant leur processus de croissance selon un cycle de 24 heures : 12 heures d’augmentation de luminosité puis 12 heures de décroissance. Contrairement aux bactéries bioluminescentes de type Vibrio, Photobacterium n’a pas besoin d’atteindre un seuil critique (quorum sensing) pour déclencher le processus de luminescence : elle émet de la lumière, même à très faible concentration. L’œil humain ne peut pas percevoir cette lumière à si faible concentration. Peut-être d’autres micro-organismes le peuvent-ils, comme les microplanctons, peut-être certains organismes plus gros. Les biologistes ne savent pas précisément pour qui cette espèce bactérienne est importante, ni pour quels usages. Ce que les biologistes savent concernant d’autres espèces de bactéries bioluminescentes, c’est qu’elles se retrouvent en général en forte concentration dans les organismes des animaux marins, soit de manière invisible ou indirectement visible, par exemple dans les tubes digestifs d’animaux, dont beaucoup ont des corps transparents, soit de manière directement visible chez les animaux qui utilisent la bioluminescence bactérienne pour leurs propres projets, par exemple à l’intérieur d’organes dédiés, les photophores. En effet, certains poissons et certains céphalopodes font de la bioluminescence par association symbiotique avec une bactérie lumineuse : ils mettent ces bactéries en culture à l’intérieur de leurs organismes et utilisent la lumière émise par la colonie afin de créer leurs propres effets de lumières grâce au système optique complexe des photophores fait de réflecteurs, d’obturateurs, de chromatophores, mais aussi de photorécepteurs qui assurent un contrôle constant et instantanée de la luminosité produite par les bactéries. De manière plus générale, il semblerait que la lumière émise par les bactéries dans le milieu sombre, profond et immense de l’océan, serve à indiquer la présence de matière organique afin d’attirer des affamés qui pourront alors héberger non-intentionnellement les bactéries dans leurs tubes digestifs. On retrouve encore ces bactéries sur les déjections, une source de nourriture pour d’autres. C’est ainsi que pourrait fonctionner Photobacterium : de tube en tube, elle coloniserait différents supports organiques emportés par le hasard des rencontres affamées. Photobacterium n’a pas été identifiée en tant que bactérie symbiotique dans le milieu marin. En labo, elle n’est pour le moment que mon associée temporaire et je ne l’ai pas encore consommée. Mais cette relation peut évoluer.
Ce sont d’abord les bactéries qu’il faut nourrir. Les souches bactériennes sont conservées au congélateur dans de petits tubes de 1 mL à -80 degrés. Elles sont réveillées et leur croissance stimulée, par inoculation dans un milieu riche. Le protocole pour la préparation de ce milieu est assez strict : un dosage précis de matière nutritive et cellulaire mélangé à de l’eau de mer, le tout rigoureusement stérilisé par autoclave avant d’être ensemencé avec la souche bactérienne sous hôte de stérilisation. En condition de laboratoire, il est prouvé qu’un milieu riche est un milieu qui crée une forte concurrence entre espèces : sans la stérilisation, il n’est pas certain que ce soit la bactérie photobacterium qui gagne la conquête du milieu. Ou plutôt, soyons précis : la stérilisation permet d’accorder tout le milieu à la seule espèce choisie par l’expérimentateur, créant ainsi une interaction organisme-milieu maîtrisé dans le but ici de produire une luminosité suffisante pour être perçue par l’œil humain. En conditions hors laboratoire, il se peut que photobacterium se développe bien, voire mieux, aux côtés d’autres micro-organismes, mais alors il se peut qu’elle ne devienne pas lumineuse, ou en tout cas pas suffisamment pour être perceptible par l’œil humain. Il se peut que ce ne soit pas du tout dans ses projets d’être perçue par l’humain·e, ceci dit, elle s’est retrouvée sur les yeux d’ANTARES de manière très visible.
En laboratoire, si le milieu n’est pas correctement stérilisé, la colonie ne produit pas de lumière. La question de savoir si photobacterium peut devenir plus lumineuse par association avec un autre organisme sous-marin reste entière. En tout cas, par association avec l’humain·e, ça fonctionne. Il semblerait ici que l’astuce biologique consistant à produire de la lumière soit un avantage évolutif pour cette espèce bactérienne à qui l’humain·e consacre tout un milieu riche en nutriments simplement pour pouvoir profiter de cette luminosité si particulière. Peut-être finira-t-elle à un moment donné par être suffisamment convaincante pour faire émerger le désir et l’idée chez un·e humain·e de la sortir de son milieu clôt, en la mangeant peut-être. À la fin du 19e siècle, certaines bactéries y étaient parvenues avec un physiologiste du nom de Raphaël Dubois qui amusait régulièrement les mondanités avec ses associations photo-bactériennes. C’est Dubois qui met au point la première recette de mise en culture de bactéries bioluminescentes, avec lesquelles il crée des lampes fournissant un éclairage étonnamment bon qu’il montre au public parisien lors de l’Exposition Universelle de 1900 aux côtés d’une autre obsession lumineuse naissante, le cinéma, et d’une nouvelle technologie émergeante, l’électricité. Quelle espèce a-t-il utilisée et qu’est-elle devenue, nul ne le sait.
Avec la bioluminescence, Dubois, auteur des premières études expérimentales et connaissances scientifiques d’envergure sur le sujet, perçoit une continuité dans l’évolution des formes vivantes à travers un « feu jamais éteint » et une continuité entre les formes vivantes par un attrait partagé pour la lumière qui nous parvient de très loin dans le temps et de très profond dans l’océan. Il nomme les deux principes actifs principaux de la bioluminescence que l’on retrouve sous différentes formes d’associations moléculaires dans toutes les espèces produisant de la lumière, la luciférine (une protéine) et la luciférase (une enzyme). Lucifer, le porteur de lumière et l’ange déchu, se retrouve au cœur d’un processus du vivant que Dubois qualifie d’ « intime », c’est-à-dire à la fois interne et propre à un corps, une création maîtrisée par un organisme dans son intérêt et dont l’intérêt se partage pourtant au-delà de lui-même, une forme donc d’autopoïese ou de sympoïese. Jean Epstein quant à lui évoque quelques années plus tard un « cinéma du diable » pour qualifier l’art de la lumière mouvante, un art animiste selon lui qui nous met en lien avec les autres formes vivantes, mais aussi les pierres, la mer, les fantômes et le temps. Peut-être sa théorie de la photogénie s’inspire-t-elle de la biophotogénie définie par Dubois grâce aux scarabées pyrophores, aux mollusques bivalves bioluminescents et aux bactéries lucifériennes. Peut-être la spécificité du cinéma, sa photogénie, n’est-elle qu’un cas particulier dans la longue histoire évolutive, la phylogenèse, de la biophotogénie. Dans ce cas, de quelle branche évolutive émerge la vision du cinéma ?
Photobacterium a été découverte sur des globes au fond de la Méditerranée. Elle est maintenant aussi présente dans des congélateurs, dans des boîtes de pétri, des tubes Falcon, et autres vaissellerie de laboratoire. J’aimerais lui redonner une visibilité sous-marine, partageable aussi par des humain·es, sans prolongements techniques. Je vais donc créer des globes transparents à l’intérieur desquels je place une colonie bactérienne afin qu’elle produise suffisamment de lumière pour faire une installation lumineuse sous-marine dans laquelle humain·es et autres organismes sous-marins locaux pourront déambuler. La bioluminescence se retrouve dans l’ensemble du monde vivant, éparpillée de manière apparemment aléatoire si l’on suit les découpages cladistiques actuels. Elle est un phénomène biologique ancien que certains datent de l’époque où apparaît la respiration, et qui a procuré dans l’océan en particulier, un avantage évolutif partagé aujourd’hui par plus de 80% de la biomasse sous-marine. La lumière attire ou repousse, signale, avertit, divertit, alerte, trompe, elle revêt des fonctions multiples et parfois contradictoires selon les connaissances scientifiques actuelles. Elle semble faire l’objet d’un certain nombre de choix de la part des organismes s’y confrontent : est-ce une lumière dont il faut s’approcher ou qu’il faut fuir ? Est-ce une lumière que je vais utiliser pour attirer, pour repousser ou pour faire diversion ? Est-ce un signal d’alliance ou de mésalliance ? La lumière bactérienne, par sa clarté douce et surtout continue, semble être plutôt attractive, indiquant une source de nourriture. Mon installation joue sur ce signalement. Humain·es et autres qu’humain·es se croiseront, tissant ainsi autrement des relations de prédations préétablies : sous l’eau et spécialement de nuit, l’humain·e n’est pas à sa place, sa fragilité est rendue sensible. Sans lumière pour éclairer les autres, c’est l’humain·e qui se retrouve éclairé·e par une autre forme de vie. Qu’est-ce que cela produira ? En quoi l’attrait de l’humain·e pour la « beauté » de cette lumière est-elle différente de l’attrait d’une seiche pour l’appétence de cette lumière ? Jusqu’où peut-on penser la prédation et où en sommes-nous de notre relation prédative aux autres espèces ? Peut-on penser les relations de l’humain·e et de la lumière avec la prédation ? La lumière est en tout cas une affaire de vie et de mort. Les bactéries le savent. Les pholades dactyles le savent. Les calmars le savent. L’humain·e le sait-iel ?
Des humain·es ancien·nes, d’il y a très longtemps, d’avant l’histoire, trouvent, avec le feu, le temps et l’espace intérieur pour raconter des histoires de leurs relations aux autres vivants : chassent, cueillettes, communications, créations, émergences, morts. Une lumière dans l’obscurité dangereuse qui rassemble et inspire des récits où commencent à apparaître quelque chose comme une individuation humaine, une conscience de son inscription singulière dans le tissu du monde. Et puis très très longtemps après, des histoires qui se racontent en lumière dans des pièces sombres, du cinéma, et puis des plus petites histoires sur de plus petits écrans, de plus en plus petits, de plus en plus attractifs, de plus en plus collés à nos peaux. L’humain·e des sociétés industrialisées utilise-t-iel la lumière ou se retrouve-t-iel domestiqué·e à l’intérieur d’une écologie lumineuse dont iel n’a pas conscience ? Le paradigme persistant de l’exception humaine au sein du vivant repose sur une image mentale fondatrice : le sujet pensant comme source de projection de la lumière sur le monde. Avec la perspective comme forme symbolique de l’inscription humaine dans le paysage s’est cristallisée une collusion entre le point de fuite, le point divin, et le point du sujet, le point de vue du Prince, l’humain idéal, absolu : l’humain en dehors du monde, dont il est ainsi maître et possesseur en tant que personnage principal et simultanément narrateur omniscient. L’univers n’est plus géocentré, mais le sensible demeure anthropocentré au travers d’un partage rigoureux, légiféré notamment par le paradigme de la propagation rectiligne de la lumière dont la source et l’horizon se confondent dans le sujet humain. Et si nous ne maîtrisions pas totalement la lumière ? Et si d’autres que nous avaient d’autres projets lumineux ? Et si nos propres projets lumineux n’étaient pas ceux que nous pensions ? Comment redéfinir une écologie dans laquelle l’humain·e est profondément entremêlé aux autres vivants avec la lumière ? Nous n’avons pas encore tout à fait déterminé les propriétés symbiotiques de l’humain·e.

Je fais passer photobacterium de tube de 1 mL en tube de 25 mL, en bocal de 1 L, en sphère de 10 L. À chaque étape d’inoculation je fais attention de ne pas introduire d’autres organismes dans le milieu afin de préparer une colonie de photobacterium la plus nombreuse et lumineuse possible. Je m’assure qu’elles aient suffisamment à manger pour 24 heures, qu’elles soient à la bonne température, et éventuellement qu’elles aient des textures intéressantes sur lesquelles s’accrocher et nicher. Autrement dit, je suis un organisme impliqué dans la croissance d’une culture bactérienne pour mes propres projets, qui, temporairement au moins, coïncident avec les projets de la bactérie, c’est-à-dire produire de la lumière pour être vue. Je repense à une relation symbiotique célèbre et souvent décrite, celle entre la sépiole euprymna scolopes et la bactérie bioluminescente vibrio fischeri. La sépiole cultive une variété spécifique de bactérie à l’intérieur de son photophore. Grâce à un système complexe de filtration et de sélection, elle parvient à trier entre l’immense quantité de bactéries du milieu afin de ne garder que celle qui l’intéresse et qui pourra lui produire de la lumière. Chaque jour elle recrache dans l’eau environnante 80% du stock bactérien usagé afin de restimuler une croissance vive. J’imagine ma sphère comme un photophore et mon protocole expérimental de stérilisation comme un processus de sélection symbiotique favorisant à la fois mon projet et celui de la bactérie sélectionnée. Dans le cas de vibrio, un phénomène très intéressant se produit chez les sépioles juvéniles qui rencontrent la bactérie pour la première fois. Le photophore de ces dernières n’est pas constitué à la naissance, il ne se forme qu’à partir de la rencontre avec vibrio, qui par un ensemble de processus de modifications génétiques « informe » ou plutôt impose une forme au photophore, afin qu’il puisse lui convenir spécifiquement. À quel moment ce type de transformation peut-il s’opérer en moi ? Existe-t-il un équivalent entre la morphogenèse de la jeune sépiole et un processus « intime » chez moi dans la rencontre avec la bactérie ?
Je rêve la nuit du système technique qui permettra aux bactéries de se développer au mieux et au public humain de voir le mieux. Je rêve des bactéries et de ce qui leur « plairait » le plus, des conditions de milieu qui leur serait le plus favorable. Je rêve des désirs de bactéries et d’un désir généralisé pour la lumière. Dans mes rêves je suis pris dans un engrenage technique, des rouages implacables, un hybride de rationalité scientifique, de sensorialité onirique, et d’une affordance ou valence non-identifiable. Comme la sépiole, mon régime d’intimité subit des transformations incontrôlées. Mais n’est-ce que la mécanique scientifique qui me submerge ou y a-t-il autre chose, quelque chose comme une morphogenèse de la pensée déclenchée par une association inter- ou plutôt trans- spécifique ? Par trans-spécifique j’insiste sur la relation de transformation mutuelle qui préside lors d’une rencontre inter-spécifique. Cependant la transformation intime qui a peut-être lieu en moi ne peut être mesurée. Elle n’est éventuellement identifiable que dans les vestiges du rêve qui transparaisse dans l’état de veille : une nouvelle idée, une image, un accident heureux. Traces d’un participio transpèce.

© Jeremie Brugidou
La bioluminescence est un phénomène relationnel. Lumière et rencontre se confondent. S’imaginer que l’humain·e serait la seule forme vivante capable d’un intérêt désintéressé pour la bioluminescence, qu’il serait absolument en dehors du phénomène et en position de contrôle neutre et objectif relève d’une abstraction épistémologique construite par la rationalité scientifique. Cette abstraction permet de produire des savoirs solides, elle permet d’identifier et de distinguer un certain nombre de phénomènes, elle permet d’éclairer le vivant d’une manière propre à l’humain·e scientifique d’éclairer, mais elle ne permet pas de nourrir la dimension relationnelle, elle ne permet pas de saisir et encore moins de décrire les transformations qui s’opèrent en nous lors de ces rencontres. Ces réflexions me viennent alors que je suis en train de construire un écosystème artificiel riche en nutriments pour les bactéries, isolé du milieu environnant dans une sphère étanche, milieu environnant dont pourtant les bactéries proviennent « naturellement ». Quelle imbrication paradoxale ! Pourtant la sphère et l’isolement racontent quelque chose de nos modes de présences : en conditions sous-marines celles-ci deviennent manifestent, mais en réalité il semblerait que l’ontologie occidentale ait produit une forme de subjectivité qui ressemble à un cerveau dans une cuve. Mon installation propose une correspondance et un possible renversement : comme ces bactéries nous sommes à la fois infiniment loin et infiniment proche du monde, entre le télescope et le microscope. Face à ces globes lumineux et bleus dans une étendue obscure où nous flottons, que devenons-nous ? Dans quelles autres visions sommes-nous projetés ?
Cet écosystème artificiel, ou cet artefact de natureculture, je le crée comme s’il s’agissait d’un photophore : la sélection à l’entrée est intraitable afin que la bactérie que j’ai choisie d’y placer profite exclusivement du milieu nutritif pour produire la plus grande intensité de lumière possible. Cette lumière je veux qu’elle pénètre la vision de l’humain·e, autrement dit, son champ d’apparition, tout en obligeant l’humain·e à s’aventurer dans d’autres situations corporelles et affectives par la plongée nocturne. Par cette double intrusion, je produis un lien avec l’océan profond d’où viennent ces bactéries, tout en manifestant la fragilité natureculturelle de ce lien par l’intermédiaire des globes clos. Il s’agit d’une machine épistémologique, moitié biologique, moitié technologique, qui vise à produire des transformations paradigmatiques chez l’humain·e à l’aide de bactéries lumineuses. L’étape suivante sera de rendre cette machine autonome, et enfin véritablement sympoïetique.
Jeremie Brugidou
L’auteur mène actuellement une recherche dans le cadre de l’IMéRA (Institut Méditerranéen de Recherches Avancées / Aix Marseille Université) en partenariat avec le MIO (Institut Méditerranéen d’Océanologie) et le Musée Subaquatique de Marseille..